
My PhD: Part 3: the return of the ichthyosaurs
In this, the final of my original trilogy describing my planned studies, I will talk about the ‘spin-offs’ from looking at the British genera and phylogeny. The other posts are:
- Part 1: a new ichthyosaur
- Part 2: the ichthyosaurs strike back
- Part 3: the return of the ichthyosaurs
This blog will look at two subsequent projects in my thesis. The first, a disparity analysis, follows from the descriptive and phylogenetic work. The second, biomechanics, follows mainly from the descriptions.
Summary
- Disparity is the study of variance
- Ichthyosaurs showed varying levels of disparity through their existence
- High disparity did not always mean high diversity
- Biomechanics is the study of animal movement
- Finite Element Analysis (FEA) is a powerful tool to study this
- Studies on skulls can tell a lot about animals’ feeding strategies
- Ichthyosaurs have yet to be studied using FEA
Disparity analysis
Disparity is the study of variation within a group of organisms. This is different from diversity, which looks at classification and systematics. Disparity uses variation to estimate differences in the ecology of an organism: how it lives in, and interacts with its environment. The two can be interlinked — greater disparity may indicate greater diversity, and _vice versa. This does not always hold true, and ichthyosaurs, being as odd as they are, show a good example of this.
Thorne, Ruta and Benton (2011) looked at ichthyosaurs over the Triassic–Jurassic boundary (TJB; ~200 Ma). Using morphospace plots — charts that show morphological variation — they showed that post-TJB ichthyosaurs had a lower disparity than pre-TJB ichthyosaurs (fig. 1). The TJB marks the spread of the thunnisaur (tuna-shaped) ichthyosaurs.
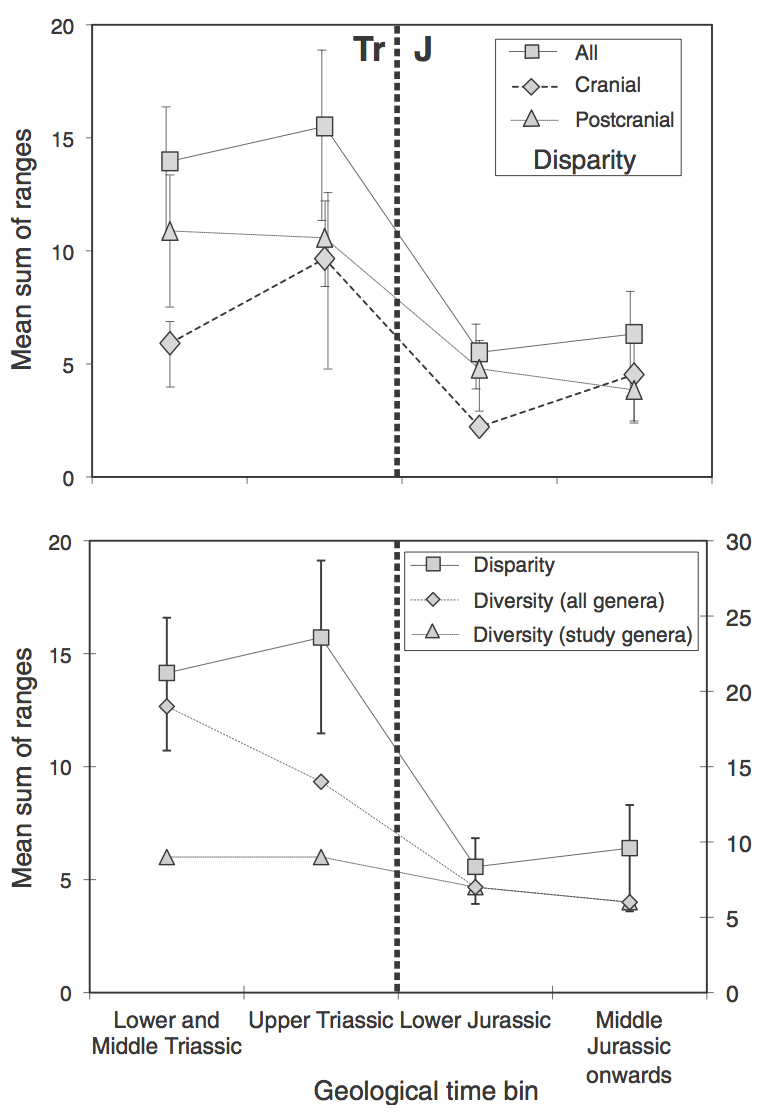
The method used in this type of study is fairly simple: measure various dimensions of the organisms. These can include length, width, height, radius, curvature etc. of individual bones, or body parts. The data are then compared using computer algorithms that assess the influence of each aspect of variation. This is usually spat out as a matrix of seemingly meaningless numbers. These numbers can then be plotted onto morphospace plots.
Usually, these morphospace plots are two-dimensional. The algorithms produce variables that show different forms of variation: e.g. longer or shorter, wider or narrower. These variables can then be plotted as principle coordinates. The two coordinates with the greatest effect or variation are most frequently chosen. The data can be further analysed to take all coordinates into account. A ‘mean sum of ranges’, for instance, is a simple way to show how great is the spread (variable).
Biomechanical studies
As I’m sure many of you will have noticed, animals have a tendency to move about. This can be by walking, running, jumping, swimming and flying. To do all of these, the animal must use its muscles and skeleton to apply forces through the feet, tail and arms. When the animal works harder more force is applied: e.g. doing a full press-up is more difficult than bending at the knees.
Using muscles and bones to apply forces, doesn’t just make you move. The bones themselves bend slightly too. The greater the forces, the more the bone is deformed. If the force is too great, the bone breaks. Studies into bone deformation are one aspect of biomechanics (Rayfield 2007).
In vertebrates, the most interesting applications of biomechanics centre on the skull. It is here where eating is done. Also, the large number of bones in the skull makes for complicated, and sometimes unexpected, interactions. As of writing, there have been no skull-biomechanics studies on ichthyosaurs (that I’ve found). My research will hopefully change that.
‘Why should you study skull biomechanics?’ I hear you cry. The skulls tell us a lot about how an animal lived. In particular, what and how it ate and how it could see. Sight in ichthyosaurs has been covered by Motani (1999). Feeding is usually compared to modern dolphins, including the schooling behaviour. Some ichthyosaur fossils have remains of belemnites — relatives of squid with a hard ‘bullet’ shell — in the stomach region.
Using biomechanics, there have been many changes in perceptions of vertebrate feeding strategies. Humphries et al. (2007) showed that pterosaurs — a group of flying reptiles, extant during the Mesozoic — could not skim feed. Skim feeding requires dipping the lower jaw into water then closing the jaws when food is caught. A pterosaur that tried to skim feed would probably break its bill!
Humphries et al. (2007) used a sequence of calculations to predict the outcome. A more complex technique used is finite element analysis (FEA). This has been used in engineering for many years. Its use in biology and palaeontology is more recent. One of the major proponents is Dr Emily Rayfield here at the University of Bristol.
FEA uses computer tomography (CT) scans of skulls as a basis model then overlays a mesh composed of an arbitrary number of fixed points. This model can then be put through computer software that applies forces to it. It location and direction of the forces is specified, as are any joins, sutures and pivots. The stress and strain experienced by the model are then calculated (fig. 2).
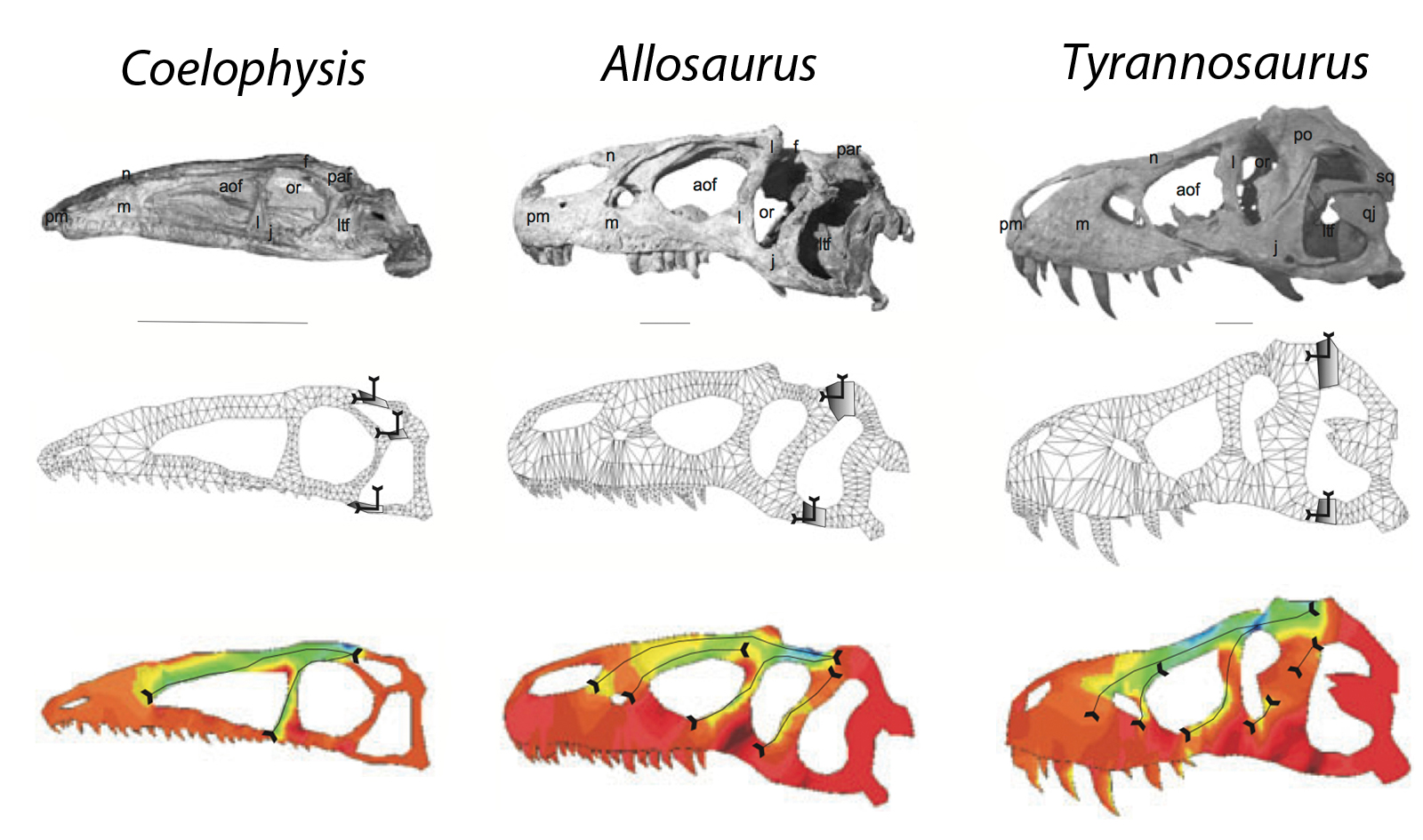
An advantage of FEA is that it is a more sophisticated technique than simpler rigid bar calculations. It takes more account of the object’s shape and the variations in strength that this causes. More accurate responses to the effects of sutures are also possible. The model and results produced are generally a more accurate representation of real life.
FEA studies have been used to great effect in studying dinosaurs (Rayfield 2004, 2005; Rayfield et al. 2007). These have been used to show that some that some ‘top predators’ (e.g. Allosaurus and Tyrannosaurus) could not use the same strategy in killing prey.
References
HUMPHRIES, S., BONSER, R. H. C., WITTON, M. P. and MARTILL, D. M. 2007. Did pterosaurs feed by skimming? Physical modelling and anatomical evaluation of an unusual feeding method. PLoS Biology, 5, 1647–1655.
MOTANI, R. 1999. Large eyeballs in diving ichthyosaurs. Nature, 402, 747.
RAYFIELD, E. J. 2005. Aspects of comparative cranial mechanics in the theropod dinosaurs Coelophysis, Allosaurus and Tyrannosaurus. Zoological Journal of the Linnean Society, 144, 309–316.
—— 2004. Cranial Mechanics and Feeding in Tyrannosaurus rex. Proceedings of the Royal Society of London Series B-Biological Sciences, 271, 1451–1459.
—— 2007. Finite Element Analysis and understanding the biomechanics and evolution of living and fossil organisms. Annual Review of Earth and Planetary Sciences, 35, 541–576.
——, MILNER, A. C., XUAN, V. B. and YOUNG, P. G. 2007. Functional Morphology of Spinosaur “Crocodile-Mimic” Dinosaurs. Journal of Vertebrate Paleontology, 27, 892–901.
THORNE, P. M., RUTA, M. and BENTON, M. J. 2011. Resetting the evolution of marine reptiles at the Triassic-Jurassic boundary. Proceedings of the National Academy of Sciences, 108, 8339–8344.
Leave a comment